



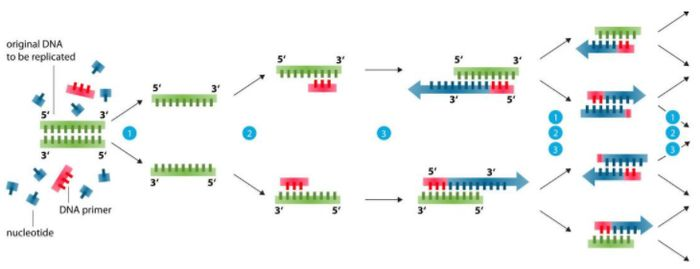
Primer design basis (99% problemer kan løses)
1. Primerlengde: Læreboken krever 15-30bp, vanligvis ca. 20bp.Den faktiske tilstanden er bedre å være 18-24bp for å sikre spesifisiteten, men jo lengre jo bedre, for lang primer vil også redusere spesifisiteten, og redusere utbyttet.
2. Primeramplifikasjonsspenn: 200-500bp er passende, og fragmentet kan utvides til 10kb under spesifikke forhold.
3. Primer base: Innholdet av G+C bør være 40-60%, for lite G+C amplifikasjonseffekt er ikke bra, for mye G+C er lett å vises uspesifikke bånd.ATGC distribueres best tilfeldig, og unngår klynger med mer enn 5 purin- eller pyrimidinnukleotider.Multi-gc for 5'-enden og mellomsekvenser for å øke stabiliteten, unngå rik GC ved 3'-enden, ingen GC for de siste 3 basene, eller ingen GC for 3 av de siste 5 basene.
4. Unngå sekundær struktur i primere, og unngå komplementering mellom to primere, spesielt komplementering i 3'-enden, ellers vil primer-dimer dannes og ikke-spesifikke amplifiserte bånd genereres.
5. Basene i 3'-enden av primerne, spesielt de siste og nest siste basene, bør være strengt paret for å unngå PCR-svikt på grunn av uparede terminale baser.
6. Primere har eller kan tilsettes med passende spaltningsseter, og den amplifiserte målsekvensen bør fortrinnsvis ha passende spaltningsseter, noe som er svært fordelaktig for spaltningsanalyse eller molekylær kloning.
7. Spesifisitet av primere: primere bør ikke ha noen åpenbar homologi med andre sekvenser i nukleinsyresekvensdatabasen.
8. Lær å bruke programvare: PP5, Oligo6, DNAstar, Vector NTI, primer3 (Dette nettdesignet fungerer best).
Innholdet ovenfor kan løse minst 99 % av primerdesignproblemene.
Kontroller detaljene i primerdesign
1. Primerlengde
Generell primerlengde er 18~30 baser.Generelt er den viktigste faktoren som bestemmer annealingstemperaturen til primeren lengden på primeren.Utglødningstemperaturen til primeren velges vanligvis (Tm-verdi -5℃), og noen bruker direkte Tm-verdi.Følgende formler kan brukes til å grovberegne annealingstemperaturen til primere.
Når lengden på primeren er mindre enn 20bp: [4(G+C)+2(A+T)]-5℃
Når lengden på primeren er større enn 20bp: 62,3℃+0,41℃(%GC)-500/lengde-5℃
I tillegg kan mange programmer også brukes til å beregne utglødningstemperaturen, beregningsprinsippet vil være annerledes, så noen ganger kan den beregnede verdien ha et lite gap.For å optimalisere PCR-reaksjoner, brukes de korteste primerne som sikrer annealingstemperaturer på ikke mindre enn 54 ℃ for best effektivitet og spesifisitet.
Totalt sett øker primerspesifisiteten med en faktor på fire for hvert ekstra nukleotid, slik at minimum primerlengde for de fleste applikasjoner er 18 nukleotider.Den øvre grensen for primerlengde er ikke veldig viktig, hovedsakelig relatert til reaksjonseffektivitet.På grunn av entropi, jo lengre primeren er, desto lavere er hastigheten som den annealer for å binde seg til mål-DNA for å danne en stabil dobbelttrådet mal for DNA-polymerase å binde.
Når du bruker programvare for å designe primere, kan lengden på primere bestemmes av TM-verdien i sin tur, spesielt for primere av fluorescens kvantitativ PCR, bør TM=60℃ eller så kontrolleres.
2.GC-innhold
Generelt er innholdet av G+C i primersekvenser 40%~60%, og GC-innhold og Tm-verdi til et primerpar bør koordineres.Hvis primeren har en alvorlig GC- eller AT-tendens, kan passende mengde A, T eller G og C hale tilsettes til 5'-enden av primeren.
3. Glødetemperatur
Utglødningstemperaturen skal være 5 ℃ lavere enn avkjettingstemperaturen.Hvis antallet primerbaser er lite, kan annealingstemperaturen økes passende, noe som kan øke spesifisiteten til PCR.Hvis antallet baser er stort, kan glødetemperaturen reduseres passende.Glødetemperaturforskjellen mellom et par primere på 4℃ ~ 6℃ vil ikke påvirke PCR-utbyttet, men ideelt sett er annealingstemperaturen til et primerpar den samme, som kan variere mellom 55℃ ~ 75℃.
4. Unngå det sekundære strukturområdet til amplifikasjonsmalen
Det er best å unngå den sekundære strukturregionen til malen når du velger det amplifiserte fragmentet.Den stabile sekundære strukturen til målfragmentet kan forutsies og estimeres av relevant dataprogramvare, noe som er nyttig for malvalg.Eksperimentelle resultater viser at ekspansjonen ofte er mislykket når den frie energien (△G) i regionen som skal utvides er mindre enn 58,6 lkJ/mol.
5. Mismatch med mål-DNA
Når den amplifiserte mål-DNA-sekvensen er stor, kan en primer binde seg til flere deler av mål-DNAet, noe som resulterer i at flere bånd vises i resultatet.Denne gangen er det nødvendig å bruke BLAST-programvaretestingen, nettstedet:http://www.ncbi.nlm.nih.gov/BLAST/.Velg Juster to sekvenser (bl2seq).
Liming av primersekvenser til sone 1 og mål-DNA-sekvenser til sone 2 kan byttes ut, og BLAST beregner komplementære, antisense og andre muligheter, slik at brukere ikke trenger å legge merke til om begge kjedene er sense-kjeder.Du kan også legge inn GI-nummeret hvis du kjenner GI-nummeret til sekvensen i databasen, slik at du ikke trenger å lime inn en stor del av sekvensen.Til slutt klikker du på Juster ved 3 for å se om primeren har flere homologe steder i mål-DNA.
6. Primerterminal
3'-enden av primeren er der forlengelsen begynner, så det er viktig å forhindre at mismatching starter der.3'-enden bør ikke være mer enn 3 påfølgende G eller C, fordi dette vil føre til at primeren feilaktig utløses i G+C-anrikningssekvensområdet.3'-enden kan ikke danne noen sekundær struktur, bortsett fra ved spesielle PCR (AS-PCR)-reaksjoner, kan ikke 3'-enden av primeren mismatches.For eksempel, hvis den kodende regionen er amplifisert, bør 3'-enden av primeren ikke termineres ved den tredje posisjonen til kodonet, fordi den tredje posisjonen til kodonet er utsatt for å degenerere, noe som vil påvirke spesifisiteten og effektiviteten til amplifikasjon.Når du bruker annekteringsprimere, se brukstabellen for kodoner, vær oppmerksom på biologiske preferanser, ikke bruk annekteringsprimere i 3′-enden, og bruk en høyere konsentrasjon av primere (1uM-3uM).
7. Sekundær struktur av primere
Primerne i seg selv bør ikke ha komplementære sekvenser, ellers vil selve primerne foldes til hårnålsstrukturer, og denne sekundære strukturen vil påvirke bindingen av primere og maler på grunn av sterisk hindring.Hvis kunstig vurdering brukes, bør de kontinuerlige komplementære basene til selve primerne ikke være større enn 3bp.Det skal ikke være noen komplementaritet mellom de to primerne, spesielt den komplementære overlappingen av 3'-enden bør unngås for å forhindre dannelsen av primer-dimere.Generelt bør det ikke være mer enn 4 påfølgende baser homologi eller komplementaritet mellom et par primere.
8. Legg til markører eller loki
5'-enden har liten effekt på amplifikasjonsspesifisiteten og kan derfor modifiseres uten å påvirke amplifikasjonsspesifisiteten.Modifikasjonen av primer 5'-enden inkluderte: tilsetning av enzymrestriksjonssete;Merket biotin, fluorescens, digoksin, Eu3+, etc. Introduser proteinbindende DNA-sekvenser;Innføring av mutasjonssteder, innsetting og manglende mutasjonssekvenser og innføring av promotorsekvenser osv. De ekstra basene vil mer eller mindre påvirke effektiviteten av amplifikasjon og øke sjansen for dannelse av primerdimer, men noen innrømmelser må gjøres for neste steg.Ytterligere sekvenser som ikke eksisterer på målsekvensen, slik som restriksjonsseter og promotersekvenser, kan legges til 5'-enden av primeren uten å påvirke spesifisiteten.Disse sekvensene er ikke inkludert i beregningen av primer Tm-verdier, men bør testes for komplementaritet og intern sekundær struktur.
9. Subkloner
Mesteparten av tiden er PCR bare foreløpig kloning, og da må vi subklone målfragmentet inn i forskjellige vektorer, så vi må designe ytterligere baser for neste operasjon i PCR-trinnet.
Noen sekvenser designet for subkloning er oppsummert nedenfor.
Restriksjonsendonuklease-restriksjonssete ble lagt til
Å legge til enzymrestriksjonssteder er den mest brukte metoden for subkloning av PCR-produkter.Generelt er spaltningsstedet seks baser, i tillegg til 5 'enden av spaltningsstedet må det legges til 2 ~ 3 beskyttende baser.Imidlertid er antallet beskyttende baser som kreves av forskjellige enzymer forskjellig.SalⅠ krever for eksempel ingen beskyttende base, EcoRⅤ krever 1 beskyttende base, NotⅠ krever 2 beskyttende baser, og Hind Ⅲ krever 3 beskyttende baser.
LIC legger til halen
Det fulle navnet til LIC er Ligation-Independent cloning, en kloningsmetode oppfunnet av Navogen spesifikt for sin del av pET-vektoren.PET-bæreren fremstilt ved LIC-metoden har ikke-komplementære 12-15 baser enkeltstrengs klebrige ender, som komplementerer de tilsvarende klebrige endene på målinnsatsfragmentet.For amplifikasjonsformål bør primer 5′-sekvensen til det innsatte fragmentet komplementere LIC-vektoren.3′→5′ ekstranect-aktiviteten til T4 DNA-polymerase kan danne en enkelt-tråd klebrig ende på det innsatte fragmentet etter kort tid.Fordi produktet kun kan dannes fra gjensidig annealing av det preparerte innskuddsfragmentet og vektoren, er denne metoden veldig rask og effektiv, og den er rettet kloning.
Rettet TA klon legge til hale
TA-kloning var ikke i stand til å målrette fragmentet inn i en vektor, så senere introduserte Invitrogen en vektor som kunne målrette kloning, som inneholdt fire fremtredende base GTGGS i den ene enden.Derfor, i utformingen av PCR-primere, bør komplementære sekvenser legges til tilsvarende, slik at fragmenter kan "orienteres".
Hvis du har kort tid, kan du prøve direkte syntese, ved å kombinere genet med vektoren, som er det vi kaller ET-gensyntese hos musekulære.
D. In-Fusion kloningsmetode
Ingen ligase nødvendig, ingen lang reaksjon nødvendig.Så lenge en sekvens i begge ender av den lineariserte vektoren er introdusert I utformingen av primere, så tilsettes PCR-produktet og den lineariserte vektoren i infusjonsenzymløsningen som inneholder BSA og plasseres ved romtemperatur i en halv time, kan transformasjonen utføres.Denne metoden er spesielt egnet for konvertering av store volum.
10. Slå sammen grunning
Noen ganger er bare begrenset sekvensinformasjon kjent om primerdesign.For eksempel, hvis bare aminosyresekvensen er kjent, kan den sammenslående primeren utformes.En fusjonsprimer er en blanding av forskjellige sekvenser som representerer alle de forskjellige basemulighetene som koder for en enkelt aminosyre.For å øke spesifisiteten kan du referere til kodonbrukstabellen for å redusere annektering i henhold til basebrukspreferanser til forskjellige organismer.Hypoxanthine kan pares med alle basene for å redusere annealingstemperaturen til primeren.Ikke bruk de annekterte basene i 3'-enden av primeren fordi annealingen av de siste 3 basene i 3'-enden er tilstrekkelig til å starte PCR på feil sted.Høyere primerkonsentrasjoner (1μM til 3μM) brukes fordi primere i mange annekteringsblandinger ikke er spesifikke for målmalen.
PCR-råvarerkontroll
1. Grunningsmengde
Konsentrasjonen av hver primer er 0,1 ~ 1 umol eller 10 ~ 100 pmol.Det er bedre å produsere det nødvendige resultatet med den laveste mengden primer.Høy konsentrasjon av primer vil forårsake mismatch og uspesifikk amplifikasjon, og øke sjansen for dannelse av dimerer mellom primere.
2. Primerkonsentrasjon
Konsentrasjonen av primere påvirker spesifisiteten.Den optimale primerkonsentrasjonen er vanligvis mellom 0,1 og 0,5μM.Høyere primerkonsentrasjoner fører til amplifisering av uspesifikke produkter.
3. Glødetemperatur for primer
En annen viktig parameter for primere er smeltetemperaturen (Tm).Dette er temperaturen når 50 % av primerne og komplementære sekvenser er representert som dobbelttrådete DNA-molekyler.Tm er nødvendig for å stille inn PCR-glødingstemperatur.Ideelt sett er annealingstemperaturen lav nok til å sikre effektiv annealing av primerne med målsekvensen, men høy nok til å redusere uspesifikk binding.Rimelig glødetemperatur fra 55 ℃ til 70 ℃.Glødetemperaturen er vanligvis satt til 5 ℃ lavere enn Tm av primeren.
Det er flere formler for innstilling av Tm, som varierer mye avhengig av formelen som brukes og sekvensen av primere.Fordi de fleste formler gir en estimert Tm-verdi, er alle utglødningstemperaturer bare et utgangspunkt.Spesifisitet kan forbedres ved å analysere flere reaksjoner som gradvis øker annealingstemperaturen.Start under estimert Tm-5 ℃, og øk gradvis utglødningstemperaturen i et trinn på 2 ℃.Høyere annealingstemperatur vil redusere dannelsen av primerdimerer og uspesifikke produkter.For best resultat bør de to primerne ha omtrentlige Tm-verdier.Hvis Tm-forskjellen til primerparene er mer enn 5 ℃, vil primerne vise en betydelig feilstart ved å bruke en lavere annealingstemperatur i syklusen.Hvis de to primerne Tm er forskjellige, setter utglødningstemperaturen til 5 ℃ lavere enn den laveste Tm.Alternativt, for å øke spesifisiteten, kan fem sykluser utføres først ved glødetemperaturer designet for høyere Tm, etterfulgt av de gjenværende syklusene ved glødetemperaturer designet for lavere Tm.Dette gjør det mulig å få en delvis kopi av destinasjonsmalen under trange forhold.
4. Primerrenhet og stabilitet
Standardrenheten til tilpassede primere er tilstrekkelig for de fleste PCR-applikasjoner.Fjerningen av benzoyl- og isobutylylgrupper ved avsalting er minimal og forstyrrer derfor ikke PCR.Noen applikasjoner krever rensing for å fjerne sekvenser som ikke er i full lengde i synteseprosessen.Disse avkortede sekvensene oppstår fordi effektiviteten til DNA-syntesekjemien ikke er 100 %.Dette er en sirkulær prosess som bruker gjentatte kjemiske reaksjoner ettersom hver base legges til for å lage DNA fra 3′ til 5′.Du kan mislykkes i begge syklusene.Lengre primere, spesielt de som er større enn 50 baser, har en stor andel trunkerte sekvenser og kan kreve rensing.
Utbyttet av primere påvirkes av effektiviteten til syntetisk kjemi og rensemetode.Biofarmasøytiske selskaper, som Cytology og Shengong, bruker alle en minimum OD-enhet for å sikre den totale produksjonen av oligonukleosid.Tilpassede primere sendes i tørr pulverform.Det er best å gjenoppløse primerne i TE slik at den endelige konsentrasjonen er 100μM.TE er bedre enn avionisert vann fordi pH-verdien i vannet ofte er sur og vil forårsake hydrolyse av oligonukleosider.
Stabiliteten til primere avhenger av lagringsforholdene.Tørt pulver og oppløste primere bør oppbevares ved -20 ℃.Primere oppløst i TE ved konsentrasjoner større enn 10 μM kan lagres stabilt ved -20 ℃ i 6 måneder, men kan bare lagres ved romtemperatur (15 ℃ til 30 ℃) i mindre enn 1 uke.Tørrpulverprimere kan lagres ved -20 C i minst 1 år og ved romtemperatur (15 C til 30 C) i opptil 2 måneder.
5. Enzymer og deres konsentrasjoner
For tiden er Taq DNA-polymerasen som brukes i utgangspunktet det genteknologiske enzymet syntetisert av koliforme bakterier.Mengden enzym som trengs for å katalysere en typisk PCR-reaksjon er ca. 2,5 U (refererer til det totale reaksjonsvolumet på 100 ul).Hvis konsentrasjonen er for høy, kan det føre til uspesifikk amplifikasjon;hvis konsentrasjonen er for lav, vil mengden syntetisk produkt reduseres.
6. Kvalitet og konsentrasjon av dNTP
Kvaliteten på dNTP er nært knyttet til konsentrasjonen og effektiviteten av PCR-amplifikasjon.dNTP-pulveret er granulært, og dets variasjon mister sin biologiske aktivitet hvis det oppbevares på feil måte.dNTP-løsningen er sur, og bør brukes i høy konsentrasjon, med 1M NaOH eller 1M Tris.HCL-bufferløsning for å justere PH til 7,0 ~ 7,5, liten mengde underemballasje, frossen lagring ved -20 ℃.Multippel fryse-tining vil degradere dNTP.Ved PCR-reaksjon bør dNTP være 50 ~ 200umol/L.Spesielt bør oppmerksomhet rettes mot at konsentrasjonen av de fire DNTPS skal være lik (lik føflekk preparat).Hvis konsentrasjonen til en av dem er forskjellig fra de andre (høyere eller lavere), vil mismatch oppstå.For lav konsentrasjon vil redusere utbyttet av PCR-produkter.dNTP kan kombineres med Mg2+ og redusere konsentrasjonen av fritt Mg2+.
7. Template (målgen) nukleinsyre
Mengden og rensegraden av templatnukleinsyre er en av nøkkelleddene for suksess eller fiasko for PCR.De tradisjonelle DNA-rensemetodene bruker vanligvis SDS og protease K for å fordøye og kaste prøver.Hovedfunksjonene til SDS er: løse opp lipider og proteiner på cellemembranen, og dermed ødelegge cellemembranen ved å løse opp membranproteiner, og dissosiere kjerneproteiner i cellen, SDS kan også kombineres med proteiner og utfelles;Protease K kan hydrolysere og fordøye proteiner, spesielt histoner bundet med DNA, og deretter bruke organisk løsningsmiddel fenol og kloroform for å trekke ut proteiner og andre cellekomponenter, og bruke etanol eller isopropylalkohol for å utfelle nukleinsyre.Den ekstraherte nukleinsyren kan brukes som en mal for PCR-reaksjoner.For generelle kliniske deteksjonsprøver kan en rask og enkel metode brukes til å løse opp celler, lysere patogener, fordøye og fjerne proteiner fra kromosomer til frie målgener, og brukes direkte til PCR-amplifisering.RNA-templateekstraksjon bruker vanligvis guanidinisotiocyanat eller protease K-metoden for å forhindre at RNase bryter ned RNA.
8.Mg2+ konsentrasjon
Mg2+ har en signifikant effekt på spesifisiteten og utbyttet av PCR-amplifikasjon.I en generell PCR-reaksjon, når konsentrasjonen av forskjellige dNTP er 200umol/L, er den passende konsentrasjonen av Mg2+ 1,5 ~ 2,0mmol/L.Mg2+-konsentrasjonen er for høy, reaksjonsspesifisiteten avtar, uspesifikk amplifikasjon forekommer, for lav konsentrasjon vil redusere aktiviteten til Taq DNA-polymerase, noe som resulterer i reduksjon av reaksjonsprodukter.
Magnesiumioner påvirker flere aspekter av PCR, slik som DNA-polymeraseaktivitet, som påvirker utbytte;Et annet eksempel er primer-annealing, som påvirker spesifisiteten.dNTP og mal binder seg til magnesiumion, og reduserer mengden fritt magnesiumion som kreves for enzymaktivitet.Den optimale magnesiumionekonsentrasjonen varierer for forskjellige primerpar og maler, men en typisk PCR-startkonsentrasjon med 200 μM dNTP er 1,5 mM (merk: For kvantitativ PCR i sanntid, bruk 3 til 5 mM magnesiumionløsning med en fluorescerende sonde).Høyere konsentrasjoner av frie magnesiumioner øker utbyttet, men øker også uspesifikk amplifikasjon og reduserer troskap.For å bestemme den optimale konsentrasjonen ble titreringer av magnesiumioner utført i trinn på 0,5 mM fra 1 mM til 3 mM.For å redusere avhengigheten av magnesiumionoptimalisering kan Platinum Taq DNA-polymerase brukes.Platina Taq DNA-polymerase er i stand til å opprettholde funksjon over et bredere spekter av magnesiumionekonsentrasjoner enn Taq DNA-polymerase og krever derfor mindre optimalisering.
9. Pcr-fremmende tilsetningsstoffer
Optimalisering av annealingstemperatur, primerdesign og magnesiumionkonsentrasjon er tilstrekkelig for svært spesifikk amplifikasjon av de fleste maler;Noen maler, inkludert de med høyt GC-innhold, krever imidlertid ytterligere tiltak.Tilsetningsstoffer som påvirker smeltetemperaturen til DNA gir en annen måte å forbedre produktspesifisitet og utbytte.Full denaturering av malen er nødvendig for best resultat.
I tillegg forhindrer den sekundære strukturen primerbinding og enzymforlengelse.
PCR-tilsetningsstoffer, inkludert formamid, DMSO, glyserin, betain og PCRx Enhancer Solution, forbedrer amplifikasjonen.Deres mulige mekanisme er å redusere smeltetemperaturen, og dermed hjelpe annealingen av primere og hjelpe DNA-polymerase-forlengelse gjennom den sekundære strukturregionen.PCRx Solution har andre fordeler.Minimal magnesiumionoptimalisering er nødvendig når den brukes med Platinum Taq DNA-polymerase og Platinum Pfx DNA-polymerase.Dermed er Platinum-teknikken kombinert med tilsetningsstoffet for å øke spesifisiteten samtidig som avhengigheten av den tredje tilnærmingen, magnesiumionoptimalisering, reduseres.For best resultat bør konsentrasjonen av tilsetningsstoffer optimaliseres, spesielt DMSO, formamid og glyserol, som hemmer Taq DNA-polymerase.

10. Varm start
Hotstart PCR er en av de viktigste metodene for å forbedre PCR-spesifisiteten i tillegg til god primerdesign.Selv om den optimale forlengelsestemperaturen til Taq DNA-polymerase er 72 ℃, forblir polymerasen aktiv ved romtemperatur.Således produseres ikke-spesifikke produkter når holdetemperaturen er lavere enn annealingstemperaturen under forberedelsen av PCR-reaksjonen og ved begynnelsen av den termiske syklusen.Når de er dannet, blir disse uspesifikke produktene effektivt forsterket.Hot-start PCR er spesielt effektiv når stedene som brukes til primerdesign er begrenset av plasseringen av genetiske elementer, slik som stedsrettede mutasjoner, ekspresjonskloning eller konstruksjon og manipulering av genetiske elementer brukt til DNA-konstruksjon.
En vanlig metode for å begrense aktiviteten til Taq DNA-polymerase er å fremstille PCR-reaksjonsløsning på is og plassere den i et forvarmet PCR-apparat.Denne metoden er enkel og rimelig, men den fullfører ikke aktiviteten til enzymet og eliminerer derfor ikke fullstendig amplifikasjonen av ikke-spesifikke produkter.
Termisk priming forsinker DNA-syntese ved å hemme en essensiell komponent til PCR-apparatet når denatureringstemperatur.De fleste manuelle termiske initieringsmetoder, inkludert forsinket tilsetning av Taq DNA-polymerase, er tungvint, spesielt for applikasjoner med høy gjennomstrømning.Andre termiske primingmetoder bruker et voksskjold for å omslutte en essensiell komponent, inkludert magnesiumioner eller enzymer, eller for å fysisk isolere reaktive komponenter, som maler og buffere.I løpet av den termiske syklusen frigjøres de ulike komponentene og blandes sammen mens voksen smelter.I likhet med den manuelle varmstartmetoden er voksskjermmetoden tungvint og utsatt for forurensning og er ikke egnet for bruk med høy gjennomstrømning.
Platina DNA-polymerase er praktisk og effektiv for automatisk varmstart-PCR.Platina Taq DNA polymerase består av rekombinant Taq DNA polymerase kombinert med monoklonalt antistoff mot Taq DNA polymerase.Antistoffene er formulert ved PCR for å hemme enzymaktivitet under langvarig temperaturholding.Taq DNA-polymerase ble frigjort til reaksjonen under 94℃-isoleringen av denatureringstrinnet, og gjenopprettet full polymeraseaktivitet.I motsetning til kjemisk modifisert Taq DNA-polymerase for termisk initiering, krever ikke platinaenzymet langvarig isolasjon ved 94 ℃ (10 til 15 minutter) for å aktivere polymerasen.Med PlatinumTaq DNA-polymerase ble 90 % av Taq DNA-polymerase-aktiviteten gjenopprettet etter 2 minutter ved 94 ℃.

11. Nest-PCR
Påfølgende runder med amplifikasjon ved bruk av nestede primere kan forbedre spesifisitet og sensitivitet.Den første runden er en standard forsterkning på 15 til 20 sykluser.En liten fraksjon av det innledende amplifikasjonsproduktet ble fortynnet 100 til 1000 ganger og tilsatt til den andre amplifikasjonsrunden i 15 til 20 sykluser.Alternativt kan det innledende amplifiserte produktet størrelsesbestemmes ved gelrensing.En nestet primer brukes i den andre amplifikasjonsrunden, som kan binde seg til målsekvensen inne i den første primeren.Bruken av nestet PCR reduserer muligheten for amplifikasjon av flere målseter fordi det er få målsekvenser som er komplementære til begge sett med primere.Det samme totale antall sykluser (30 til 40) med de samme primerne amplifiserte de uspesifikke setene.Nested PCR øker sensitiviteten til begrensede målsekvenser (f.eks. sjeldne mrnas) og forbedrer spesifisiteten til vanskelig PCRS (f.eks. 5′ RACE).
12. Synkende PCR
Synkende PCR forbedrer spesifisiteten ved å bruke tette annealingsforhold for de første par PCR-syklusene.Syklusen starter ved en glødetemperatur som er omtrent 5 ℃ høyere enn estimert Tm, deretter reduseres hver syklus med 1 ℃ til 2 ℃ til glødetemperaturen er under Tm 5 ℃.Bare destinasjonsmalen med høyest homologi vil bli forsterket.Disse produktene fortsetter å ekspandere i påfølgende sykluser, og fortrenger de forsterkede uspesifikke produktene.Synkende PCR er nyttig for metoder der graden av homologi mellom primer og målmal ikke er kjent, for eksempel AFLP DNA-fingeravtrykk.
Relaterte PCR-sett
2× PCR-heltenTMMix-systemet har høyere toleranse for PCR-hemmere enn det ordinære PCR Mix-systemet, og kan enkelt takle PCR-amplifisering av ulike komplekse maler.Det unike reaksjonssystemet og høyeffektive Taq Hero gjør at PCR-reaksjonen har høyere amplifikasjonseffektivitet, spesifisitet og sensitivitet.
Høyere forsterkningseffektivitet
Den har 5'→3' DNA-polymeraseaktivitet og 5'→3' eksonukleaseaktivitet, uten 3'→5' eksonukleaseaktivitet.
Real Time PCR Easyᵀᴹ-SYBR Green I Kit
Spesifikt – optimalisert buffer og hot-start Taq-enzym kan forhindre uspesifikk amplifikasjon og dannelse av primerdimer
Høy følsomhet – kan oppdage lave kopier av malen
Settet bruker et unikt Foregene revers transkripsjonsreagens og Foregene HotStar Taq DNA-polymerase kombinert med et unikt reaksjonssystem for å effektivt forbedre amplifikasjonseffektiviteten og spesifisiteten til reaksjonen.
Innleggstid: mai-09-2023



