



一、Øk reaksjonssystemets følsomhet:
1. Isoler RNA av høy kvalitet:
Vellykket cDNA-syntese kommer fra RNA av høy kvalitet.RNA av høy kvalitet bør minst være i full lengde og fri for revers transkriptasehemmere som EDTA eller SDS.Kvaliteten på RNA bestemmer den maksimale mengden sekvensinformasjon du kan transkribere til cDNA.En vanlig RNA-rensemetode er en ett-trinns metode som bruker guanidin isotiocyanat/syrefenol.For å forhindre kontaminering med spormengder av RNase, må RNA isolert fra RNase-rike prøver (som bukspyttkjertel) lagres i formaldehyd for å bevare høykvalitets RNA, spesielt for langtidslagring.RNA ekstrahert fra rottelever ble i hovedsak nedbrutt etter å ha vært lagret i vann i en uke, mens RNA ekstrahert fra rottemilt holdt seg stabilt etter å ha vært lagret i vann i 3 år.I tillegg er transkripsjoner lengre enn 4 kb mer følsomme for nedbrytning av spor-RNaser enn små transkripsjoner.For å øke stabiliteten til lagrede RNA-prøver, kan RNA oppløses i avionisert formamid og lagres ved -70 °C.Formamid som brukes til å konservere RNA må være fritt for RNA-nedbrytende rusk.RNA fra bukspyttkjertelen kan bevares i formamid i minst ett år.Når du forbereder å bruke RNA, kan du bruke følgende metode for å utfelle RNA: tilsett NaCl til 0,2M og 4 ganger volumet av etanol, plasser ved romtemperatur i 3-5 minutter og sentrifuger ved 10 000×g i 5 minutter.
2. Bruk den RNaseH-inaktive (RNaseH-) revers transkriptase:
RNase-hemmere legges ofte til revers transkripsjonsreaksjoner for å øke lengden og utbyttet av cDNA-syntese.RNase-hemmere bør tilsettes under den første trådsyntesereaksjonen i nærvær av en buffer og et reduksjonsmiddel (som DTT), fordi prosessen før cDNA-syntese denaturerer inhibitoren, og frigjør dermed bundet RNase som kan bryte ned RNA.Protein RNase-hemmere forhindrer kun nedbrytning av RNA av RNase A, B, C, og forhindrer ikke RNase på huden, så vær forsiktig så du ikke introduserer RNase fra fingrene til tross for bruk av disse inhibitorene.
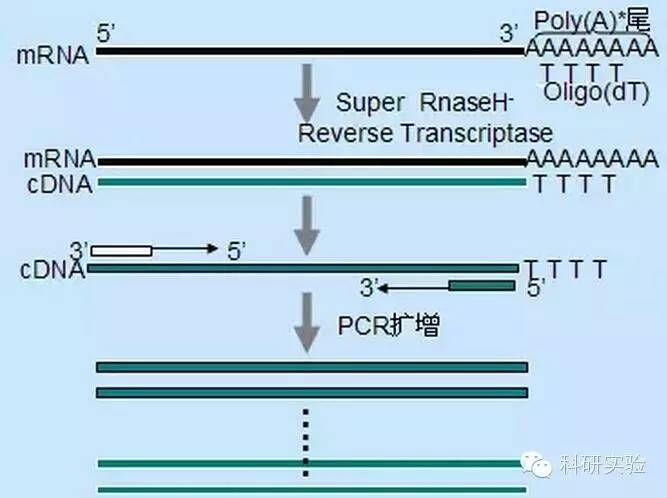
Revers transkriptase katalyserer omdannelsen av RNA til cDNA.Både M-MLV og AMV har endogen RNaseH-aktivitet i tillegg til sin egen polymeraseaktivitet.RNaseH-aktivitet og polymeraseaktivitet konkurrerer med hverandre om hybridtråden som dannes mellom RNA-malen og DNA-primeren eller cDNA-forlengelsesstrengen, og bryter ned RNA-tråden i RNA:DNA-komplekset.RNA-malen degradert av RNaseH-aktivitet kan ikke lenger tjene som et effektivt substrat for cDNA-syntese, noe som reduserer utbyttet og lengden av cDNA-syntesen.Derfor vil det være fordelaktig å eliminere eller sterkt redusere RNaseH-aktiviteten til revers transkriptase.
SuperScript Ⅱ revers transkriptase, RNaseH-MMLV revers transkriptase og thermoScript revers transkriptase, RNaseH-AMV, kan oppnå mer mengde og mer full-lengde cDNA enn MMLV og AMV.RT-PCR-sensitivitet vil bli påvirket av mengden cDNA-syntese.ThermoScript er mye mer følsomt enn AMV.Størrelsen på RT-PCR-produkter er begrenset av revers transkriptases evne til å syntetisere cDNA, spesielt ved kloning av større cDNA.Sammenlignet med MMLV, økte SuperScripⅡ betydelig utbyttet av lange RT-PCR-produkter.RNaseH-revers transkriptase har også økt termostabilitet, slik at reaksjonen kan utføres ved temperaturer høyere enn de normale 37-42°C.Under de foreslåtte syntesebetingelsene, bruk oligo(dT)-primer og 10 μCi av [α-P]dCTP.Det totale utbyttet av første tråd ble beregnet ved å bruke TCA-utfellingsmetoden.Full-lengde cDNA ble analysert ved bruk av størrelsessorterte bånd som ble skåret ut og talt på en alkalisk agarosegel.
3. Øk inkubasjonstemperaturen for omvendt transkripsjon:
En høyere inkubasjonstemperatur bidrar til å åpne den sekundære RNA-strukturen, og øker reaksjonsutbyttet.For de fleste RNA-maler vil inkubering av RNA og primere ved 65 °C uten buffer eller salt, etterfulgt av rask avkjøling på is, eliminere de fleste sekundære strukturer og tillate primere å binde seg.Imidlertid har noen maler fortsatt sekundære strukturer, selv etter varmedenaturering.Amplifisering av disse vanskelige malene kan utføres ved å bruke ThermoScript Revers Transcriptase og plassere revers transkripsjonsreaksjonen ved en høyere temperatur for å forbedre amplifikasjonen.Høyere inkubasjonstemperaturer kan også øke spesifisiteten, spesielt når genspesifikke primere (GSP) brukes til cDNA-syntese (se kapittel 3).Hvis du bruker GSP, sørg for at Tm til primerne er den samme som forventet inkubasjonstemperatur.Ikke bruk oligo(dT) og tilfeldige primere over 60°C.Tilfeldige primere krever inkubasjon ved 25°C i 10 minutter før de øker til 60°C.I tillegg til å bruke en høyere revers transkripsjonstemperatur, kan spesifisiteten også forbedres ved direkte å overføre RNA/primer-blandingen fra 65 °C denatureringstemperaturen til revers transkripsjonsinkubasjonstemperaturen og tilsette en forhåndsoppvarmet 2× reaksjonsblanding (cDNA varmstartsyntese).Denne tilnærmingen bidrar til å forhindre den intermolekylære baseparingen som oppstår ved lavere temperaturer.Den multiple temperaturvekslingen som kreves for RT-PCR kan forenkles ved å bruke en termisk syklus.
Den termostabile polymerase virker som en DNA-polymerase i nærvær av Mg2+ og som en RNA-polymerase i nærvær av Mn2+.Den kan holdes varm ved en maksimal temperatur på 65°C.Tilstedeværelsen av Mn2+ under PCR reduserer imidlertid troskap, noe som gjør Tth-polymerase mindre egnet for høypresisjonsforsterkning, slik som kloning av cDNA.I tillegg har Tth lav revers transkripsjonseffektivitet, noe som reduserer følsomheten, og siden revers transkripsjon og PCR kan utføres med et enkelt enzym, kan ikke kontrollreaksjoner uten revers transkripsjon brukes til å sammenligne cDNA-amplifikasjonsprodukter med kontaminerende genomisk DNA.Amplifikasjonsproduktene ble separert.
4. Tilsetningsstoffer som fremmer omvendt transkripsjon:
Tilsetningsstoffer inkludert glyserol og DMSO tilsettes første-strengs syntesereaksjonen, noe som kan redusere stabiliteten til nukleinsyredobbeltstrengen og løsne den sekundære strukturen til RNA.Opptil 20 % glyserol eller 10 % DMSO kan tilsettes uten å påvirke SuperScript II- eller MMLV-aktivitet.AMV kan også tolerere opptil 20 % glyserol uten tap av aktivitet.For å maksimere sensitiviteten til RT-PCR i SuperScriptⅡ revers transkripsjonsreaksjon, kan 10 % glyserol tilsettes og inkuberes ved 45 °C.Hvis 1/10 av revers transkripsjonsreaksjonsproduktet tilsettes PCR, er konsentrasjonen av glyserol i amplifikasjonsreaksjonen 0,4 %, noe som ikke er nok til å hemme PCR.
5. RNaseH-behandling:
Behandling av cDNA-syntesereaksjoner med RNaseH før PCR kan øke følsomheten.For noen maler antas det at RNA i cDNA-syntesereaksjonen forhindrer binding av amplifikasjonsprodukter, i så fall kan RNaseH-behandling øke følsomheten.Generelt er RNaseH-behandling nødvendig ved amplifisering av lengre cDNA-målmaler i full lengde, slik som tuberøs scherosis II med lav kopi.For denne vanskelige malen forbedret RNaseH-behandling signalet produsert av SuperScript II eller AMV-syntetisert cDNA.For de fleste RT-PCR-reaksjoner er RNaseH-behandling valgfri, fordi PCR-denatureringstrinnet ved 95 °C generelt hydrolyserer RNA i RNA:DNA-komplekset.
6. Forbedring av deteksjonsmetode for små RNA:
RT-PCR er spesielt utfordrende når bare små mengder RNA er tilgjengelig.Glykogen tilsatt som en bærer under RNA-isolering bidrar til å øke utbyttet av små prøver.RNase-fritt glykogen kan tilsettes samtidig som Trizol tilsettes.Glykogen er vannløselig og kan holdes i den vandige fasen med RNA for å hjelpe påfølgende utfelling.For prøver mindre enn 50 mg vev eller 106 dyrkede celler er den anbefalte konsentrasjonen av RNase-fritt glykogen 250 μg/ml.
Tilsetning av acetylert BSA til den omvendte transkripsjonsreaksjonen ved bruk av SuperScript II kan øke følsomheten, og for små mengder RNA kan reduksjon av mengden SuperScript II og tilsetning av 40 enheter RNaseOut nukleasehemmer øke deteksjonsnivået.Hvis glykogen brukes i RNA-isolasjonsprosessen, anbefales det fortsatt å legge til BSA eller RNase-hemmer ved bruk av SuperScript II for revers transkripsjonsreaksjon.
二、Øk RT-PCR-spesifisiteten
1. CND-asyntese:
Første-strengs cDNA-syntese kan initieres ved hjelp av tre forskjellige metoder, hvis relative spesifisitet påvirker mengden og typen av syntetisert cDNA.
Den tilfeldige primermetoden var den minst spesifikke av de tre metodene.Primere annealerer på flere steder gjennom transkripsjonen, og genererer korte cDNA-er med delvis lengde.Denne metoden brukes ofte for å oppnå 5′-endesekvenser og for å oppnå cDNA fra RNA-maler med regioner med sekundær struktur eller med termineringssteder som ikke kan replikeres med revers transkriptase.For å oppnå det lengste cDNA, må forholdet mellom primere og RNA i hver RNA-prøve bestemmes empirisk.Startkonsentrasjonen av tilfeldige primere varierte fra 50 til 250 ng per 20 μl reaksjon.Siden cDNA syntetisert fra totalt RNA ved bruk av tilfeldige primere primært er ribosomalt RNA, velges generelt poly(A)+RNA som mal.
Oligo(dT)-primere er mer spesifikke enn tilfeldige primere.Den hybridiserer til poly(A)-halen som finnes i 3'-enden av de fleste eukaryote mRNA-er.Fordi poly(A)+ RNA er omtrent 1 % til 2 % av totalt RNA, er mengden og kompleksiteten til cDNA mye mindre enn med tilfeldige primere.På grunn av sin høye spesifisitet krever oligo(dT) generelt ikke optimalisering av forholdet mellom RNA og primere og poly(A)+-seleksjon.Det anbefales å bruke 0,5 μg oligo(dT) per 20 μl reaksjonssystem.oligo(dT)12-18 er egnet for de fleste RT-PCR.ThermoScript RT-PCR-systemet tilbyr oligo(dT)20 på grunn av dets bedre termiske stabilitet for høyere inkubasjonstemperaturer.
Genspesifikke primere (GSP) er de mest spesifikke primerne for revers transkripsjonstrinnet.GSP er et antisense-oligonukleotid som spesifikt kan hybridisere til RNA-målsekvensen, i motsetning til tilfeldige primere eller oligo(dT), som annealer til alle RNA-er.De samme reglene som brukes til å designe PCR-primere gjelder for utformingen av GSP i revers transkripsjonsreaksjoner.GSP-en kan være den samme sekvensen som amplifikasjonsprimeren som annealer til den 3′-endeste enden av mRNA, eller GSP-en kan utformes for å anneale nedstrøms for revers amplifikasjonsprimeren.For noen amplifiserte forsøkspersoner må mer enn én antisense-primer utformes for vellykket RT-PCR fordi den sekundære strukturen til mål-RNA kan forhindre primerbinding.Det anbefales å bruke 1 pmol antisense GSP i en 20 μl førstestrengs syntesereaksjon.
2. Øk inkubasjonstemperaturen for omvendt transkripsjon:
For å dra full nytte av den fulle fordelen av GSP-spesifisitet, bør en revers transkriptase med høyere termostabilitet brukes.Termostabile revers transkriptaser kan inkuberes ved høyere temperaturer for å øke reaksjonsstringensen.For eksempel, hvis en GSP annealer ved 55°C, vil ikke spesifisiteten til GSP bli utnyttet fullt ut hvis AMV eller M-MLV brukes for revers transkripsjon ved en lav stringens på 37°C.SuperScript II og ThermoScript kan imidlertid reageres ved 50°C eller høyere, noe som vil eliminere uspesifikke produkter generert ved lavere temperaturer.For maksimal spesifisitet kan RNA/primer-blandingen overføres direkte fra denatureringstemperaturen på 65°C til inkubasjonstemperaturen for revers transkripsjon og tilsettes til en forvarmet 2x reaksjonsblanding (cDNA-syntese varmstart).Dette bidrar til å forhindre intermolekylær baseparing ved lave temperaturer.De flere temperaturovergangene som kreves for RT-PCR kan forenkles ved å bruke en termisk syklus.
3. Reduserer genomisk DNA-forurensning:
En potensiell vanskelighet man møter med RT-PCR er forurensning av genomisk DNA i RNA.Ved å bruke en god RNA-isoleringsmetode, for eksempel Trizol Reagent, vil mengden av genomisk DNA som forurenser RNA-preparatet reduseres.For å unngå produkter avledet fra genomisk DNA, kan RNA behandles med amplifikasjonsgrad DNase I for å fjerne kontaminerende DNA før revers transkripsjon.DNase I-fordøyelsen ble avsluttet ved å inkubere prøvene i 2,0 mM EDTA i 10 minutter ved 65°C.EDTA kan chelatere magnesiumioner, og forhindrer magnesiumionavhengig RNA-hydrolyse ved høye temperaturer.
For å skille amplifisert cDNA fra kontaminerende genomiske DNA-amplifikasjonsprodukter, kan primere designes som hver annealer til separate eksoner.PCR-produkter avledet fra cDNA vil være kortere enn de avledet fra kontaminert genomisk DNA.I tillegg ble et kontrolleksperiment uten revers transkripsjon utført på hver RNA-mal for å bestemme om et gitt fragment var avledet fra genomisk DNA eller cDNA.PCR-produktet oppnådd uten revers transkripsjon er avledet fra genomet.
Innleggstid: 16. mai 2023



